Fas���ڰ�Ѫ��ϸ��HL-60��Stat3�ı�������ữ
�����ڶ���ҽ��ѧѧ��2000���21����6��
�����桡���̿�������Ƽ��������
����ժ��Ҫ��Ŀ��:�۲�Fas��HL-60ϸ�����쳣��ֳ�Ƿ��е������á�����:�û������鼼����Fas��ϸ��������Ĥ����IL-6�źŴ�����gp130ϸ����������Ƕ�������壨Fas/130����ͬʱ����Fas��������FasDD���滻Fas/130��130ϸ�������ṹ�֣�Fas/130f�����ֱ���HL-60�б�����ÿ�Fas���弤����ЩǶ�������壬����������黯�͵�����ӡ������������ϸ�������γ�ͬԴ�������壨130cyt-130cyt-130cyt��FasDD-FasDD-FasDD����ϸ����Stat3�ı��P���ữ�� ���:תȾpED fas/130���ÿ�Fas��������6��8 h��HL-60ϸ��Stat3�����½�(P��0.05)����Fas/130f���Stat3�ı����½�������(P��0.01)����ʵ������Fas/130f���������Կ����յ�10 minʱ,Stat3 Tyr705���ữ��Fas/130�����Խ���(P��0.01)�����ۣ�Fas���������ư�Ѫ��HL-60ϸ����Stat3�ı���,��ͬʱ����Stat3 Tyr705���ữ��
�����ؼ��ʣ�ϸ�����ӣ����壻��Ѫ�����������źŴ���
������Ѫ��ϸ�����쳣��ֳ��Ӱ���ϸ���������ܵ���Ҫԭ���������쳣��ֳ��Ŀǰ��һ���о��ȵ㡣��Ѫ���������ӣ�LIF�������ư�Ѫ��ϸ������ֳ��1����������ЧӦ���������ϰ�ϸ��Ĥ�ϵ�LIF������ǻ�(gp190)����ʹ֮����һ�ǻ���gp130���γ���Դ�Զ�������2�����Ӷ�ʹϸ����һϵ���źŷ��ӷ����仯��3����Ȼ����LIF�����÷dz��㷺���ѱ��ų��ں�ѡ����ҩ��֮�⡣Ŀǰ����ȷ�� fas��һ�������������Եĵ������ӣ�����ϸ��������һ���鵼������Ϣ�Ľṹ��FasDD����4�������о��û������鼼���Ʊ�Ƕ�������壨Fas/130�� Fas/130f��,���ֱ���HL-60�б����������ϸ����Stat3�ı��P���ữˮƽ,��ȷ��Fas�������Ѫ��ϸ����ֳ��Ӱ�졣
����1�����Ϻͷ���
����1.1��ϸ���ꡡHL-60ϸ�������п�Ժ�Ϻ�ϸ�����ṩ����ϸ�����ú�8%СţѪ��� dMEM ��Gibco��37�棬5% CO2 ������������
����1.2��FasǶ������Ʊ�����pRclgo gp130��Moreau��ʿ�ṩ��Ϊģ�壬�ֱ���Sal��/Xho ��,Xba ��ø�в�װ��pALTER������Ϊ��5��-GTCTCGCTTT cTAGAGCAGAACAGCAC-3�䣬����Altered Sites ��ͻ��ϵͳ��Promega��ʹ��Ĥ����ϸ������֮�����Xba��ø��λ�㡣ͬ����pBluescript�� Fas (Taupin ��ʿ����) ��ͬ���������� Xba ��(��� 5��- C tTC CTT TCT AGA CAC CCA AAC AA- 3��) ����Hind ���Xba �� ø�з���Fasϸ�������Ϳ�Ĥ������gp130ϸ��������130cyt)����ΪFas/130�������Hind ��/Sal��EcoR ����Ƕ�����pALTER������������pED��Moreau��ʿ�ṩ����
����1.3��Fas�����ṹ��ķ����װ�䡡��PCR����Fas������Ƭ�Σ�N��(5��-GT AAG GAT CCC ACC ACT ATT GC-3��) �� C�� (5��-TTC TGG ATC CCT AGT AAT gTC C-3��) ����һBamH ��λ�㡣Ȼ�� ��Bgl��-Bgl��ȥ��gp130ϸ�������ĵ�66��153������(88 AA),����Fas�ĵ� 233��320������ȡ���������ɺ�FasDD��Ƕ���� fas/130f��
����1.4��FasǶ��������HL-60ϸ��תȾ������Ƕ�������ķ�ת¼��������pED��pED fas/130 ��pED Fas/130f����֬����תȾ����Gibco-BRL��˾��LipofectamineTM�Ͳ������裩�ֱ�תȾ��HL-60��72 h���ռ�ϸ�����á�
����1.5�������黯���Stat3�ı��תȾ�ɹ����ϸ���ú�200 ng/ml��Fas�������������������8��12 h���������䶳PBS��ϸ��ϴ2�Σ����Ž���4%��ȩ�й̶�30 min������Ϳ���ڸ��и������������ز�Ƭ�ϣ�37���ҹ�����ǰ��Ϳ��ϸ�����ز�Ƭ���¹̶�20 min��Ȼ����PBSϴ3�Ρ��μ�������Ѫ����20 min���ÿ���Stat3 igG(Pharmagene)37�淴Ӧ1 h����AKP�������IgG(Promega)��ͬ����������ϸ���������������ڼ�������ø����Һ��pH9.5��5 min������ BCIP/NBTʹ֮��ɫ�������۲�ϸ����ɫ�ʵ���������ˮ��ϴ����ɡ�����������Ƭ��������ͼ�����ϵͳ�����գ�FR-988 smartScape2000������ǡ�������
����1.6�������黯���Stat3�����ữ��תȾϸ������Ѫ������������24 h���뿹Fas ���巴Ӧ0��30 min,��������4%��ȩ�̶���һ��Ϊ�ÿ������ữStat3��Tyr705������������ͬ�ϡ�
����1.7��������ӡ������Stat3�ı�����䶳PBS��ϸ��ϴ2�Σ�Ȼ��5��106��ϸ����100��l TBS��pH 7.4, 20 mmol/L Tris-HCl��150 mmol/L NaCl��1 mmol/L EDTA������1% triton X-100 200 ��mol/L ��PMSF��Sigma�� 2 ��mol/L ��Leupeptin 2 ��mol/L aprotinine����˳�Լ���˾�����ڱ��Ϸ�Ӧ20 min��,4������ 12 000 r/min 20 min��������12% SDS-PAGE������Ӿ���ò�ʽתĤ�ǣ�TE52X pharmacia Biotech��������ת�Ƶ�PVDF��Whatman�����ÿ���Stat3 IgG��Ĥ����ϣ�AKP��ǵĿ���IgG��һ����ϣ�BCIP/NBT��Ϊø������ɫ��
����2���ᡡ��
����2.1��FasǶ������Ʊ�����Altered Sites ��ͻ��ϵͳ�������gp130�Ŀ�Ĥ����ϸ�����������������������øλ�㾭Xba��ø��֤ʵ��gp130ϸ������Fas��ϸ��������Ĥ�������Լ�FasDD������������ȫ�Զ�DNA�����ǣ�ABI pRISM 373A����������ȷ����
����2.2��FasǶ��������HL-60ϸ��תȾ����֬���彫pED Fas/130 ��pED fas/130f�ֱ�תȾ��HL-60ϸ������֤ʵ��Ƕ����ֱ���HL-60ϸ���б�����5������
����2.3�������黯���Stat3�ı������200��500 ng/ml ����Fas���壬��Fas/130��Fas/130f�е�Fasϸ��������϶��ֱ��γ������壬�۲����ϸ����Stat3����ˮƽ���������,Stat3����Fas/130��϶�����Ϊ�ͣ�P��0.05������Fas/130f��ȴ��Fas/130���ͣ�P��0.01,ͼ1����ʵ��֤����130ϸ�������γ�������ʱ������Stat3�ı��Fas������Ҳ��Эͬ���á����⣬�����Ʊ���һ190/130fǶ������6����Ϊ���գ���ȷ��Fas�������ڵ���ʱ�Ƿ��л��ԡ�������֣�190/130f��ϸ���ڵ�Stat3ˮƽ��հ��������ͼ2��������Fas�������������ϸ����,��������ЧӦ��
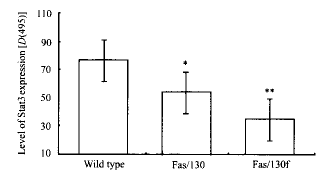
ͼ 1����Ƕ����HL-60��Stat3ˮƽ
����fig 1��Stat3 expression in HL-60 transfected
����with Fas/130 or Fas/130f
����P��0.05; ��������P��0.01 vs wild type
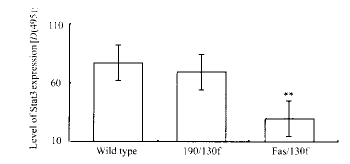
ͼ 2��Fas��������Ѿ�����Stat3�Ĺ�ϵ
����fig 2��Stat3 level in HL-60 transfected
����with 190/130f of Fas/130f��������P��0.01 vs wild type
����2��4�������黯����Stat3���ữ�ı仯����Fas���巴Ӧ0,5,10,15,20,30 min ��ʵ�����ʱ����Stat3���ữˮƽ���϶�����Ϊ�ͣ���Fas/130f����10 minʱStat3���ữˮƽ��Fas/130Ϊ�ͣ�ͼ3����ʵ�������Fas�������Stat3�����ữ��Ӱ�졣
����2��5��������ӡ������Stat3�ı����ͼ4���Կ����� Fas/130���Fas/130f���Stat3�ı���϶�����͡�
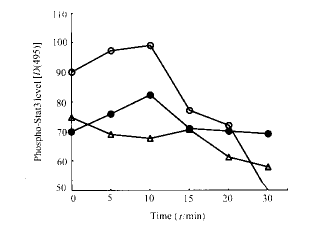
ͼ 3��Fas�������HL-60��Stat3���ữ��Ӱ��
����fig 3��Various levels of phospho-Stat3
����in the groups�� Wile type ; �� Fas/130; ���� Fas/130f
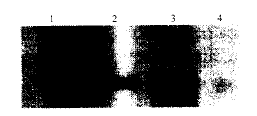
ͼ 4����Fas�����յ�Fas/130��Fas/130f�Ѿۻ���ϸ����Stat3�ı仯
����fig 4��Stat3 level in HL-60 transfected with Fas/130
����or Fas/130f after anti-Fas IgG induction
����1�� The group of Fas/130f; 2�� The group of Fas/130
����3�� HL-60 with pED�� 4�� HL-60 without anti-Fas IgG
����3���֡���
����Fas��һ�����������Ե�ϸ���������ӣ�Ϊ319��������Ŀ�Ĥ�ǵ��ס������ܰ�ϸ��������ϸ������Ƥϸ���ȱ��Fas���壨Fas l���Ϳ�Fas������Fasϸ��������Ϻ���ϸ�������ĵ��������ṹ�����仯���Ӷ�ϸ�������е�Fas���������η��Ӷ��ٽ�ϸ��������7������
����TNF ��������Ա����Fas�����������崫����Ϣ����7����Ʃ�磬 Fas L��7�� �Ϳ�Fas������8���յ�Fas���������ź�����Fas����������ʽ�����ֵġ�����G-CSF����G-CSF/FasǶ�����Fasϸ�������γɵĶ�����ȴ���ܲ���ЧӦ��9�������о���Ƶ�Ƕ����ϸ������ΪFas������������ gp130ϸ��������Fas��������Fas�����յ��γ������壬����Fas��������ϸ�����źŷ��ӵĹ�ϵ��
����Fas �յ�ϸ�������ķ���ָ���в��٣�������֤ʵFas�Ĺ��ܰ���������ֳϸ�����Ŀ�ԭ��PCNA���ı����c-myc�ĺϳ���10����������ȷ,��caspases����֬ø����֪��3��MAPK;���� nF-kappa B �Ȳ�����Fas��TNF������źŴ�����11����Ȼ����Fas��������ϸ���������������������Ȼ���������ǵ�ʵ�������Fas�������γ��������HL-60ϸ���ڵ�Stat3���Լ��٣�ͼ1����ͬʱ���ڿ�Fas��������10 minʱ��Fas/130f���Stat3 Tyr705 �����ữ��Fas/130��Ҳ�����½���ͼ3�������Զ϶���Fas������鵼��ϸ�������źŰ�������Stat3�ı��������Stat3�����ữ������ϸ����Stat3�ĺ����½����ԣ�˵��Fas������Ӱ��Stat3������ѧ����һ���������źŷ��ӵĽ��롣
������������Ŀ��������Ȼ��ѧ����������Ŀ(39670683)
������������������棨1957�������У����壩�������ڣ�˶ʿ����ʦ�����ߵ�λ�������棨�ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
�������̿����ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
��������Ƽ���ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433)
���������٣��ڶ���ҽ��ѧ����ҽѧ����֯��̥ѧ�����ң��Ϻ�200433��
���������
������1�ݡ�Metcalf d. The leukemia inhibitory factor��J��. Intern J cell cloning, 1991��9(2):95-108.
������2�ݡ�Gearing DP, Thut CJ, VandaBos T, et al. Leukemia inhibitory factor receptor is structurally related to the IL-6 signal transducer gp130��J��. eMBO J,1991,10(10):2839-2848.
������3�ݡ������棬������.��Ѫ����������������ϸ�����źŷ��ӣ�J��.����ҽѧ*��������ѧ�ֲᣬ1998,20(6):166-169.
������4�ݡ�Rieux-Laucat F, Le Deist F, Roberts AG, et al. Mutations in Fas associated with human lymphoproliferative syndrome and autoimmunity��J��. science, 1995, 268(5215):1347-1349.
������5�ݡ������棬���̿�������Ƽ,��. Fas�����ṹ����ڰ�Ѫ��ϸ�� meg-01 ��c-myc�ı����J��. �л�ѪҺѧ��־��2000(������).
������6�ݡ������棬����Ƽ��������,��.��Ѫ����������������gp130ϸ�������յ���Ѫ��ϸ��Meg-01��c-myc�ı����J��.�ڶ���ҽ��ѧѧ����2000,21(6):522-525.
������7�ݡ�Tanaka M, Suda T, Takahashi T, et al. Expression of the functional soluble form of human Fas ligand in activated lymphocyte��J��. EMBO J�� 1995,14(6): 1129-1135.
������8�ݡ�Dhein J, Daniel PT, Trauth BC,et al. Induction of apoptosis by monoclonal antibody anti-APO-1 class switch variants independent on cross-linking of APO-1 cell surface antigens��J��. J Immunol, 1992, 149(10):3166-3173.
������9�ݡ�Takahashi T, Tanaka M, Ogasawara J, et al. Swapping between Fas and granulocyte colony-stimulating factor receptor��J��. J Biol Chem, 1996,271(29): 17555-17560.
������10�� Peng X�� Maruo T�� Matsuo H, et al. Serum deprivation-induced apoptosis in cultured porcine granulosa cells is characterized by increased expression of p53 protein, Fas antigen and Fas ligand and by decreased expression of PCNA��J��. Endocr J��1998, 45(2): 247-253.
������11�� Wallach D, Varfolomeev EE, Malinin NL, et al. Tumor necrosis factor receptor and Fas signaling mechanisms��J��. Annu Rev Immunol, 1999, 17:331-346.