组成核酸的元素有C、H、O、N、P等,其中N含量约为15%~16%,磷含量为9%~10%。由于核酸分子中的磷含量比较恒定,因此,核酸定量测定的经典方法,是以测定磷含量代表核酸量。
核酸经水解可得到多核苷核,因此核苷酸是核酸的基本单位,核酸就是由很多单核苷酸聚合形成的多核苷酸,核苷酸可被水解产生核苷和磷酸,核苷还可进一步水解,产生戊糖和含氮碱。因此,核酸是由含氮碱、戊糖及磷酸三种成分组成。
含氮碱(简称碱基):核酸中的含氮碱简称碱基,是嘌呤碱(purine)与嘧啶碱(pyrimidine)的衍生物。RNA和DNA含有的共同碱基成分是腺嘌呤(adenine,A)、鸟嘌呤(guanine,G)和胞嘧啶(cytosine, C)。二者的区别是RNA含有尿嘧啶(uracil,U),而DNA含有胸腺嘧啶(thymine,T)。嘌呤和嘧啶都有酮-烯醇式互变异构现象,一般生理pH条件下呈酮式。它们的结构如下:
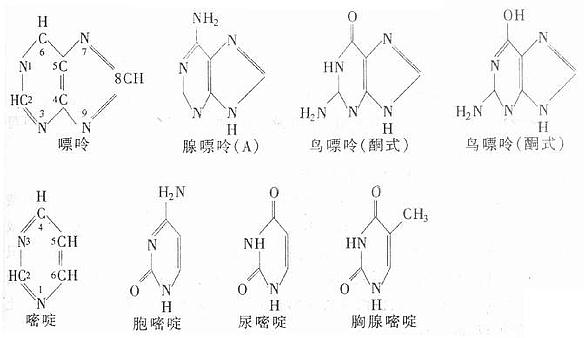
有些核酸中含有修饰碱基(或稀有碱基),这些碱基大多是在上述嘌呤或嘧啶碱的不同部位甲基化(methylation)或进行其它的化学修饰而形成的衍生物。例如有些DNA分子中含有5-甲基胞嘧啶(m5C),5-羟甲基胞嘧啶(hm5C)。某些RNA分子中含有1-甲基腺嘌呤(m1A)、2,2-二甲基鸟嘌呤(m22G)和5,6-二氢尿嘧啶(DHU)等。
嘌呤碱和嘧啶碱一般多不易溶于水,对250~280nm波长的紫外光有较强的吸收,但对260nm光波的吸收能力最大。由于碱基是核酸的基本组成成分,因此,所有的核酸(包括DNA和RNA)其共同特点是对260nm处的紫外光有最大的吸收值。
核酸分子中碱基的克分子数与磷的克原子数相等,所以可根据核酸溶液中的磷含量及紫外光的吸收值来测定核酸量。一般以每升核酸溶液中含1g磷原子为标准来计算核酸的吸光率,这称为克原子磷吸光率或克原子磷消光系数[ε(p)],ε(p)的计算式为:
ε(p)=A/Cl
A为吸光度(光密度),1为比色杯内径,通常为1.0cm;C为每升核酸溶液中磷的克原子数。
C= 每升溶液中磷重(wg)/30.98ε(p)=30.98A/wl
一般DNA的ε(p)=6000~8000;RNA为7000~10000
现将DNA和RNA的化学组成归纳如表18-1:
表18-1
| RNA | DNA | |
| 嘌呤碱 | 腺嘌呤鸟嘌呤 | 腺嘌呤鸟嘌呤 |
| 嘧啶碱 | 胞嘧啶尿嘧啶 | 胞嘧啶胸腺嘧啶(或5-甲基胞嘧啶) |
| 戊糖 | 核糖 | 脱氧核糖 |
| 磷酸 | 磷酸 | 磷酸 |
核酸又称多核苷酸,组成DNA的脱氧核糖核苷酸主要为四种,即dAMP、dGMP、dCMP及dTMP;组成RNA的核糖核苷酸主要有AMP、GMP、CMP及UMP四种。对两种核酸的组成可简写如下:
DNA=(碱基-脱氧核糖-磷酸)n;RNA=(碱基-核糖-磷酸)n
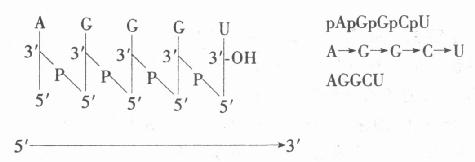
核酸中核苷酸的连接方式为:一个核苷核C-3’上羟基与下一个核苷核酸C-5’连接在磷酸羟基脱水缩合成酯键,称酯键称3’5’磷酸二酯键,若干个核苷酸间依3’5’磷酸二酯键连接成长链的大分子即为核酸。此长链称多核苷酸链,在链的一核苷酸,其C-5’连接的磷酸只一个酯键,称此核苷酸为链的5’磷酸未端或5’。链的另一端核苷酸上C-3’上羟基是自由的,对此核苷酸称为3’羟基末端或3’端,链内的核苷酸在C-5’上磷酸已形成二酯键,C-3’上羟基也已参与二酯键的形成,故称核苷酸残基。
核酸的一级结构乃指其核苷酸链中核苷酸的排列顺序,由于核酸中核苷酸彼此之间的差别乃在于碱基部分,故核酸的一级结构即指核酸分子中碱基的排列顺序。
对核酸一级结构的描述为:将5’磷酸末端书于左侧,中间部分为核苷酸残基,3’羟基末端书于右侧。通常用竖线表示核糖,碱基标于竖线上端,竖线间有含P的斜线,代表3’,5’磷酸二酯键。此表示法及简化式如下:
核酸的多核苷酸链在次级键的基础上,还可形成更为复杂的二级及三级的高级结构。
1.二级结构1953年Watson 及Crick在化学分析及X光衍射法观察DNA结构的基础上提出了著名的DNA双螺旋结构模型(double helix model)此结构是在核酸一级结构基础上形成的更为复杂的高级结构,即DNA的二级结构,结构如图18-1。
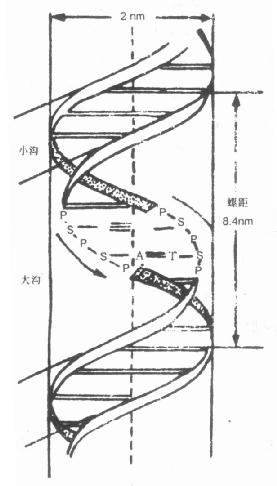
图18-1 DNA的双螺旋结构
P:磷酸基;S:脱氧核糖;G:鸟嘌呤;
A:腺嘌呤;T:胸腺嘧啶C:胞嘧啶
DNA的二级结构即双螺旋结构,其内容可归纳为:
(1)DNA分子为二条多核苷酸链以一共同轴为中心,盘绕成右手双螺旋结构。螺旋直径2nm。螺旋盘绕形成链间的两种沟,即宽的大沟与狭窄的小沟。
(2)二条多核苷酸链的走向相反,通常取左侧链从上到下为5’→3’端,右则链从下向上为5’→3’端,这样二条链构成反平行排列的双螺旋。
(3)二条多核苷酸链借氢键而连系在一起。氢键乃一链碱基上-NH2的氢与另一链上碱基的氧或氮形成。碱基有二个氢键,G与C之间有三个氢键(图18-3)。这种相配关系称为碱基互补或碱基配对。配对的碱基处于同一平面,此平面与双螺旋的中心轴垂直,由于二条链中碱基互补,所以二链彼此又称为互补链。
(4)碱基对之间氢键的能量为3~7kcal/mol,由于氢键多,所以可维系DNA双链结构。另外碱基对彼此间距离为0.34nm,每一螺旋含10个碱基对,故螺距为3.4nm,相邻碱基对间彼此尚有范德瓦士(van der Waals)力作用(此力量为1~2kcal/mol,作用范围为0.5nm),能量虽弱但由于碱基对多,合力也就大。可见碱基对的氢键及碱基对之间的范德瓦士力是稳定DNA成双螺旋结构的主要能量。
上述DNA的双螺旋结构是溶液及活体中常见的形式,通称B型。当B型所处条件的湿度低于75%时,可转变为A型。A型的碱基对不垂直于双螺旋的轴、倾斜约20度,螺距降为2.8nm,每一螺旋含11个碱基对。B型与A型的水合程度不同,它们是DNA分子在天然条件下的两种基本形式。除A、B型外尚发出有C型双螺旋,似B型,螺距3.3nm,第一螺旋含9个碱基对。
DNA双螺旋结构阐明的量重要意义在于第一次提出了遗传信息是以DNA分子中核苷酸的排列顺序为储存方式,从而说明了天然遗传信息的复制过程。
2.三级结构已发现线粒体、叶绿体、细菌、质粒及一些病毒的DNA双螺旋分子尚可形成封闭环状,天然状态的环状DNA分子多扭曲成麻花状的超螺旋结构(superhelix),这些比螺旋更为复杂的结构即DNA分子的三级结构。
真核生理细胞核中的DNA具有一种超螺旋结构,即DNA双螺旋盘绕在组蛋白上形成核小体(nucleosome)。核小体是染色质(chromatin)的核心小粒,由有140个碱基对的双螺旋DNA缠绕于由组蛋白(H2A、H2B、H3及H4各二分子)组成的八聚体外面,这一DNA股由此形成直径为9nm的超螺旋1.75圈。此核小体又经60个碱基对的DNA双螺旋及组蛋白H1形成细丝(间隔区)与下一个核小体相连接。
核小体的DNA双螺旋为200个碱基对,长度应为0.34nm×200=68nm,但实际长度只10nm,说明DNA双螺旋链进一步螺旋化盘绕在组蛋白八聚体上,其长度压缩了7/8。每6个核小体又绕成一圈形成螺线管,外径为30nm,螺距为10nm。这样DNA分子长度被压缩了6/7。120个螺线管又盘绕成直径为400nm,高为30nm的超螺线管,DNA分子长度又被压缩了40/41,此超螺线管即染色体的单位纤维(unit fiber),长20~60nm。从单位纤维形成染色单体(chromatid),实际长度为2~10nm,DNA分子长度又被压缩了5/6~6/7。这样从许多核小体组成的串珠样纤维经多层次螺旋化结构到形成染色单体,DNA分子的长度已被压缩至近1/10000。
RNA分子也是由核苷酸依3’,5’磷酸二酯键形成的多核苷酸链。RNA总是以单链的形式存在,也有5’磷末端及3’羟基末端。RNA单链的局部折叠成的某一片段的A及G分别与另一片段的U及C配对常形成发夹结构(hairpin structure)。在此结构内的碱基无需全部配对,而配对部位形成小的双螺旋区域,不能配对的碱基则连成小环从螺旋区中被圈出来。这种RNA单链局部小双螺旋结构即是RNA的二级结构。
由于DNA分子中核苷酸序列分析以及一级结构与功能相关的研究,使得人们有可能进一步了解DNA一级结构与基因组织的关系。已经证实,自然界极大多数生物体遗传信息贮存在DNA的核苷酸排列顺序中,因此,基因是DNA的一个片段,只有少数病毒的遗传信息贮存在RNA分子中。DNA分子中不同区域有不同功能,有些区域可编码蛋白质(最终产物是蛋白质),有些区域可编码RNA(最终产物是tRNA和rRNA),有些序列则与调控有关。那么一个DNA分子能携带多少基因呢?如果以平均1000个碱基对可编码一个3kD的蛋白质计算,猴病毒(SV40)DNA分子量为3.0×105,有5000碱基对可编码5种蛋白质。人染色体DNA有2.3×109碱基对,可编码200万以上的基因,但实际上,最多可编码基因数为2~3万。这是由于真核细胞DNA分子除编码蛋白质和RNA等结构基因外,有相当部分DNA顺序属于非编码区,而原核细胞DNA由于分子较小,必需充分利用有限的核苷酸序列。各种生物体内DNA分子的大小见表18-2。
表18-2 各种生物体内DNA分子的大小
| 来源 | 分子量 | 碱基对数目 | 长度 |
| 噬菌体фX174 | 0.6μm | ||
| 腺病毒(SV40) | 1.6×106 | 4500 | 1.5μm |
| 鼠线粒体 | 3.0×106 | 14000 | 4.9μm |
| 噬菌体λ | 9.5×106 | 50000 | 17μm |
| 噬菌体T2或T4 | 3.3×107 | 200000 | 67μm |
| 大肠杆菌染色体 | 1.3×108 | 4500000 | 1.5μm |
| 人染色体 | 3.0×109 | 125000000 | 4.1μm |
1.真核生物的基因组织根据某一段核苷酸顺序在整个DNA分子中出现的频率不同可分为以下几种:
(1)单拷贝顺序(singlecopy sequence):在整个DNA分子中只出现一次或少数几次,主要是编码蛋白质的结构基因。除组蛋白、角蛋白和肌动蛋白以外,几乎所有的蛋白质基因都是单拷贝顺序,平均为1000碱基对。单拷贝基因在整个基因组织中所占比例最高。在人的细胞中约占DNA含量的一半。
(2)中等重复顺序(moderatelyrepetitive sequences):有些基因如核蛋白体RNA基因、tRNA基因、组蛋白基因等在DNA分子中可重复出现几十到几千次,约占人细胞DNA含量的30~40%。以rRNA为例,在大肠杆菌中重复频率为7而果蝇中可重复千次。可见真核细胞中重复顺序比原核细胞高得多。
(3)高重复顺序(highlyrepetitive sequences):可重复几百万次。往往是简单的重复顺序,如蟹的T-A-T-A-T-A-T。也有的较长如非洲绿猴DNA是以172个碱基对的顺序为基础重复几万次。高重复顺序一般位于异染色质上,多数不编码蛋白质或RNA,其功能还不太清楚,主要是起间隔作用,可能与调控有关。
在重复顺序中还有一种反转重复顺序(inverted repetitivesequences)。其特点是一段碱基呈现回文结构,即一条单链回折即可形成互补的双链,故称为回文结构(palindromicstructure)或发夹结构(hairpin structure)。这种结构对基因的复制与转录可能具有调节 控制功能。

真核细胞中单拷贝顺序和重复顺序常常是中间隔排列的。不仅如此,在一个基因内部往往被一个或几个额外的顺序分割成若干片段,这种插入到基因内部的顺序称为插入顺序或内含子(intron)。内含子是不编码的顺序,而编码的碱基顺序则称为外显子(exon)。插入顺序是真核细胞DNA最主要的特征。
真核生物由于存在着较多的重复顺序、特殊的插入顺序以及控制区和其它多余顺序,使得DNA总长度往往大于编码的结构基因,因此,实际基因数往往小于DNA分子。
2.原核生物的基因组织原核生物DNA分子较小,基因组织也较简单,一般具有以下特点:
(1)DNA分子绝大部分用于编码蛋白质,不编码部分(又称间隔区)通常包含控制基因表达的顺序。例如,噬菌体фX174中只有5%是非编码区。
(2)功能相关的基因常常串联在一起,并转录在同一个mRNA分子中,称为多顺序反子。这种现象在真核生物中是很少见的。
(3)基因重叠:例如фX174的E基因全部包括在D基因内,B基因则包括在查基因内。这种现象主要发现在病理DNA分子中,可能是由于DNA分子太小又要装入相当量的基因的缘故。
| Copyright @ 2002-2010 婵犵數濮烽弫鍛婃叏閻戣棄鏋侀柟闂寸绾惧鏌i幇顒佹儓闁搞劌鍊块弻娑㈩敃閿濆棛顦ョ紓浣哄Т缂嶅﹪寮诲澶婁紶闁告洦鍓欏▍锝夋⒑缁嬭儻顫﹂柛鏃€鍨垮濠氬Χ閸氥倕婀遍埀顒婄秵閸嬪懘鎮甸幒妤佲拺缂備焦锚缁楁帡鏌ㄩ弴銊ら偗鐎殿喖顭烽弫鎰板川閸屾稒顥堥柛鈹惧亾濡炪倖甯掔€氼參宕戠€n喚鍙撻柛銉e妿閳藉鏌i幘瀵哥疄闁哄矉绻濆畷姗€鏁愰崨顒€顥氶梻鍌欑窔濞佳兠洪妶鍥e亾濮橆偄宓嗛柣娑卞枛椤粓鍩€椤掑嫨鈧礁鈽夊Ο婊勬閸┾偓妞ゆ帊鑳堕々鏌ユ煕閹炬せ鍋撻柛瀣尵閹叉挳宕熼鍌ゆК婵犵數鍋涢惇浼村垂閽樺鏆﹂柟杈剧畱缁犳稒銇勯弬鍨挃闁挎稒绻堝铏圭矙閹稿孩鎷遍梺鑽ゅ暀閸ヤ礁娲弫鍌涙叏閹邦亞鐩庨梻浣瑰濡礁螞閸曨垼鏁佹繝濠傜墛閻撴瑧绱掑☉姗嗗剰濞存粎澧楅幈銊︾節閸愨斂浠㈤悗瑙勬处閸嬪﹤鐣烽悢纰辨晣闁绘ǹ浜粈鍐⒒閸屾瑧顦﹂柟鑺ョ矋閹便劑鎮界粙璺唶闂佽鍨奸悘娑㈡晲婢跺﹦鐤€闂佸搫顦冲▔鏇㈡儓閸曨垱鈷戦柛鎾村絻娴滅偤鏌涢悩鍐插闁宠绉规慨鈧柕鍫濇閹锋椽鏌i悩鍏呰埅闁告柨鑻埢宥夊箛閻楀牏鍘甸梺鍛婂灟閸婃牜鈧熬鎷�. xxmy.com 闂傚倸鍊搁崐鎼佸磹閹间礁纾归柣鎴eГ閸ゅ嫰鏌涢幘鑼槮闁搞劍绻冮妵鍕冀椤愵澀鏉梺閫炲苯澧柛鐔告綑閻g兘濡歌閸嬫挸鈽夊▍顓т簼缁傚秵娼忛妸褏鐦堥梺姹囧灲濞佳冪摥闂備胶枪閿曘倝顢氶鐘愁潟闁圭偓鍓氬ḿ鈺呮煠閸濄儲鏆╅柛妯哄船椤啴濡堕崱妤€顫庢繝娈垮枟閹稿啿鐣峰┑鍡╂僵閺夊牃鏅濋敍婊堟⒑缂佹﹫渚涢柛鐘崇墵瀹曟繈鏁冮崒娑氬幐闁诲繒鍋熼弲顐㈡毄婵$偑浼囬崒婊呯崲闂佸搫鏈惄顖炵嵁濡吋宕夐柣鎴炨缚閳ь剝顕ч—鍐Χ閸℃ḿ鈹涚紓鍌氱С缁舵岸鐛崘鈺冾浄閻庯綆浜滅粣娑欑節閻㈤潧孝闁哥噥鍨堕、鏃堟偄閸忓皷鎷绘繛杈剧秬婵倗娑甸崼鏇熺厱闁挎繂绻掗悾鍨殽閻愯尙绠婚柡浣规崌閺佹捇鏁撻敓锟� 濠电姷鏁告慨鐑藉极閸涘﹥鍙忛柣鎴f閺嬩線鏌涘☉姗堟敾闁告瑥绻戦妵鍕箻閸楃偟浠肩紒鐐劤椤兘寮婚悢鐓庣鐟滃繒寮ч埀顒傜磽娴g瓔鍤欐俊顐g箞瀵寮撮姀鐘诲敹濠电娀娼уú銈壦囬埡鍛拺闁硅偐鍋涢埀顒佺墪鍗遍柛顐g妇閺€浠嬫煟濡櫣浠涢柡鍡忔櫅閳规垿顢欑拠鎻掔ギ閻庤娲濋~澶岀紦娴犲绀堥柛娆忣槹濞呮捇姊绘担鍛婅础濠⒀勵殔椤灝螣閼测晙绗夊銈嗙墱閸嬬偤鎮¢妷鈺傜厽闁哄洨鍋涢埀顒€婀遍埀顒佺啲閹凤拷10017704闂傚倸鍊搁崐鎼佸磹閹间礁纾瑰瀣捣閻棗銆掑锝呬壕濡ょ姷鍋涢ˇ鐢稿极閹剧粯鍋愰柛鎰紦閻㈠姊婚崒娆戣窗闁告挻鐟х划鏃傗偓闈涙憸娑撳秵銇勯弽顐沪闁绘挶鍎甸弻锝夊即閻愭祴鍋撻崷顓涘亾濮樼偓瀚� |