为了解释别构酶协同效应的机制并推导出动力学曲线的方程式,不少人曾提出过各种模式,各有优缺点,现在主要把Hill模式叙述如下:
Hill模式
在协同结合模式中最早的一种是Hill在1909年提出的,企图解释氧结合至血红蛋白的S形饱和曲线,现称为Hill模式,后来经Atkinson应用于别构酶反应,他设想在这个系统中,n分子的配体(S)能够一步结合到酶上去:
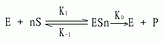
即此反应的总解离常数(K's)由下式决定
| K'S=[E][S]n/[ESS] | (6-7) |
而酶的饱和分数
| YS=每分子酶蛋白上已结合的底物分子数/每分子酶蛋白上底物结合位点的总数 | (6-8) |
又因总的酶浓度[E0]=[E]+[ES0]
| 故 YS=[ESn]/[E0]=[ESn]/[ESn]+[E] | (6-9) |
合并式6-7和式6-9,消去[ESn],则
| YS=[S]n/K'S+[S]n | (6-10) | ||
| YSK'S+YS[S]n=[S]n, | |||
| YSK'S=(1-Y)[S]n | (6-11) | ||
| Ys/1-Ys=[S]n/K'S | (6-11) | ||
| log | YS | =nlog[S]-logK'S | (6-13) |
| 1-YS | |||
因此以
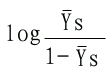
因v=k0[ESu],Vm=k0[E0],故
 =[ESu]/[E0]=v/vω =[ESu]/[E0]=v/vω | (6-14) |
将式6-10代入式6-14,即得
| [S]n/K'S+[S]n=v/Vm | |
| Vm[S]n=K'Sv+v[S]n | (6-15) |
| (Vm-v)[S]n=K'Sv | (6-16) |
| v/Vm-v=[S]n/K'S | (6-17) |
| logv/Vm-v=nlog[S]-logK'S | (6-18) |
式6-13或6-18即为Hill方程式,式6-18如以logv/Vm-v对log[S]作图,也可得一直线(见下图)。
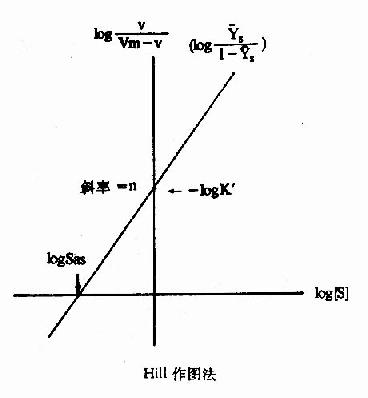
Hill作图法
如v=Vm/2时,式6-19为log1=nlog[S]-logK'S=0(6-19)
此时的[S]即S0.5,故nlogS0.5=logK'5(6-20)式6-18所得的直线斜率为n,纵轴截距为-logk'S,而横轴截距为logK'S/n,即log[S]0.5,但[S0.5]也可在已知logK'S后通过式6?0求取。
上节已述及,S0-5就相当于米曼氏动力学中的Km,当K0《k-1/k1时,可反映别构酶对底物的亲和力,S0.5愈小,亲和力愈大,而K's实际上已与亲和力关系不大,因受到n的影响。故反映底物亲和力的参数,已从非别构酶的Km一项移到别构酶的[S]一项,并且式6?0可看出K'S是随[S]而改变的,不是一个常数。由于K'S的测定是假设V=(1/2)Vm或[S]=S0.5的条件下计算的,故有些作者用S0.5S,来代表别构酶的K'5,以免与Km混淆。
Hill作图法的斜率n,称为Hill系数,即前述的协同系数,一般可用nH或h代表。当nH=1时,式6-1变为V=Vm[S]/(K1+[S]),即米曼氏方程式,表示无协同作用,此时K'或S0.5S,=S0-5=Km,至于nH>1为正协同,nH<1为负协同。
Hill模式比较简单,式6-1或式6-10都是S形曲线方程式,但有不少缺点:(1)按理,Hill系数应等于酶分子中可能有结合底物的位点数,但因忽略了ESn-1,ESn-2…ES1等中间形式的酶底物复合体,根据Hill氏作图计算出来的n值一般均低于真实的位点数。以别构蛋白Hb为例,理论上每分子Hb可结合四分子氧,即n=4,但计算结果n=2.6~2.8。在负协同效应时,每分子酶也结合n个底物(n>1)但计算结果却是n<1。故Hill系数已不能代表结合底物的位点数,而只能作为底物协同性的指标。(2)在S浓度过高(酶90%以上被S饱和)或过低(酶仅10%以下被S饱和)时,Hill线的斜率n常等于1,故当测定别构酶活力时,[S]的范围较广,得出的Hill线不是直线而是折线(见下图)。(3)n分子的底物同时和酶作用,反应的级数为n+1,如n-4则为五级反应,这在动力学上是不可能的。但尽管如此,Hill作图法仍不失是一个求取别构酶S0.5和鉴定协同类型及协同作用大小的常用方法。
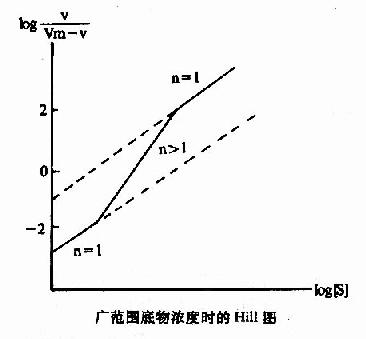
广范围底物浓度时的Hill图
| Copyright @ 2002-2010 婵炴垶鎼╅崢鑲╀焊閻楀牏鈻旀い鎾偓宕囥偐缂備礁顦拌摫闁哄瞼鍠撶槐鏃堟晸閿燂拷. xxmy.com 闂佺粯顨呴悧濠傖缚閸儱绠ラ柍褜鍓熷鐢告晸閿燂拷 濠电姭鍋撴俊銈囨嚕P婵犮垼顔愰幏锟�10017704闂佸憡鐟遍幏锟� |